


Reef-er madness
The multiple faces of the coral reef

Announcement: post too long for email!
Ecosystems are funny things. We tend to treat them as real, tangible places, something that we can touch, see, and walk through if need be. “That place there is a forest, that’s an ecosystem”. “And there, that pond is an ecosystem”. But really, ecosystems are more like a conceptual fractal of context-dependent interactions between different species of organisms, some of which are partially dictated by immutable climatic and geological considerations.
They scale all the way from covering areas the size of countries (tundras, various types of forests, deserts) to places as small as the surface of a leaf, or a thimbleful of water. As long as a given environment is such that certain species will preferentially live there (or only live there) and co-occur with other species with the same limitations, an ecosystem will arise. Conversely, since the species are the basis of the ecosystem, an area with the exact same basic conditions could give rise to radically different ecosystems depending on which species are present there (ecological succession, invasive species, etc).

Despite involving dozens, hundreds, or even thousands of species, ecosystems can be fragile things. Certain species have an outsized influence on the characteristics of the habitat, so that removing them completely alters the fabric of the community. Consider a stream, which, with a beaver, might become a pond where newts and lotic plants might live, but would just be a regular stretch of water without the animal and its dam. These organisms are known as “ecosystem engineers”. These typically aren’t apex predators, or species that directly interact with many others (see: “keystone species”), but they are species that change the characteristics of the environment to suit their needs, or through the physical space they take, create their own environment. Asian clams, for example, are ecosystem engineers whose prodigious water filtration rate changes the turbidity and plankton content of bodies of water, changing what fish and birds will be found there. 1
Throughout the course of geological time climate changes, new continents form and break-up, and species come and go. Some ecological niches have remained extremely consistent over the course of Earth’s history, while others are surprisingly new. For example, the first trees, and thus the first forests, date from the Devonian, some 390 million years ago. While the tallest species of the time didn’t seem to grow more than a dozen metres tall, they had all the bells and whistles of modern trees (except for flowers, but we’ll get back to that).2 Some species from that time, such as tree-ferns, adopted forms that can still be found today in tropical and subtropical regions of the world.

Conversely, some ecological niches are (comparatively) recent. The first flowering plants are about 140 million years old3 (although it’s hard to say for sure) and the first proto-pollinating insects (regardless of how or why they were covered in pollen back then) are a few million years younger 4. Imagine a period of nearly 250 million years that had luxurious forests, but without a single flower anywhere in sight, or any grasslands! Entire new ecosystems, new ways for species to interact with each other, arising over the course of aeons.
This dynamic is not unique to the land. Although many ecological niches have been created and filled for far longer in the sea, since life first arose there, the system is still dynamic enough that species come and go in specific roles, or create variations for themselves. One of the most interesting, and in fact most ancient, is the niche we call “the reef builders”.
Generalities regarding modern coral reefs
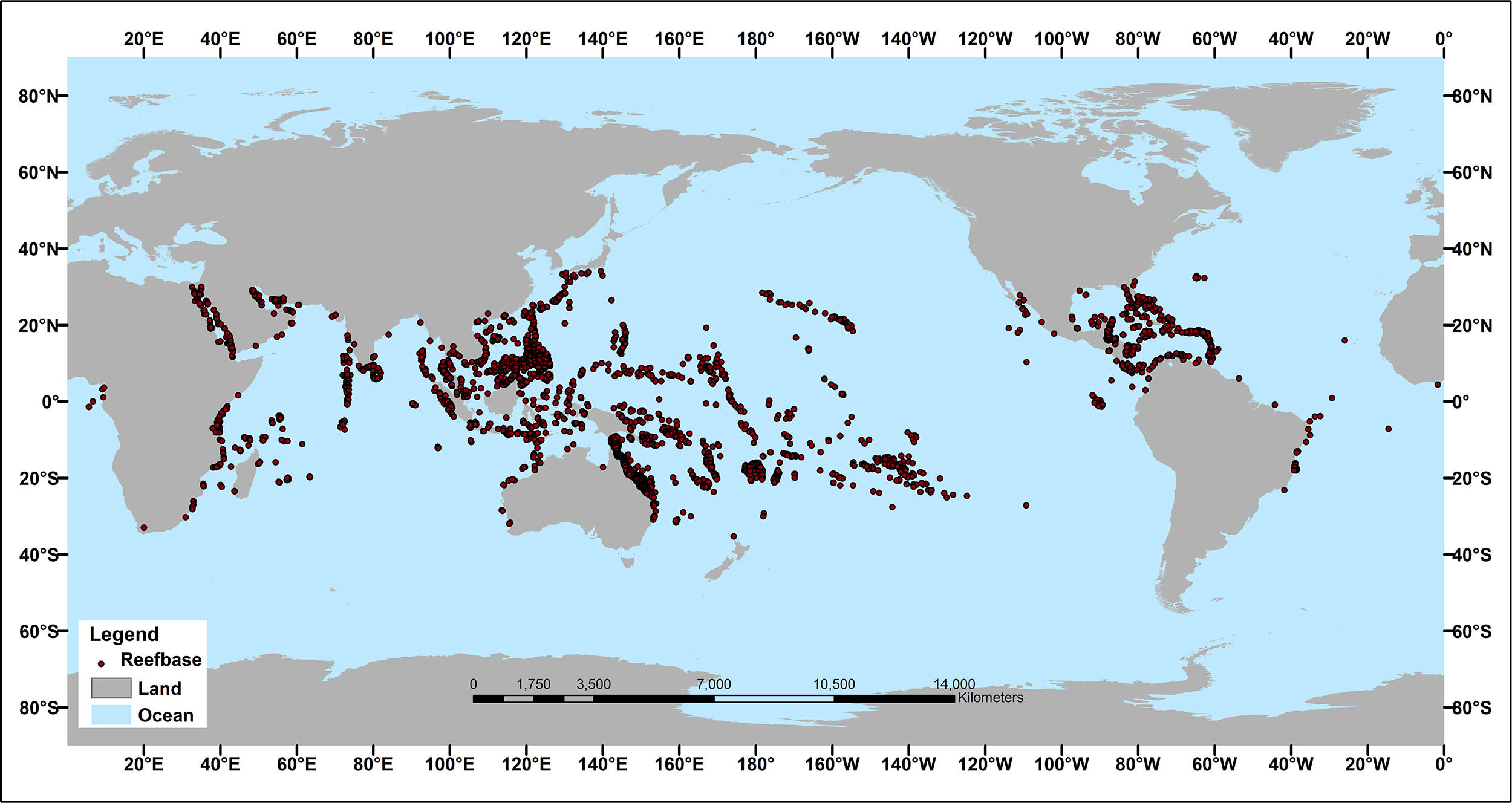
Modern coral reefs are hardened aragonite (calcium carbonate) structures generally made from the hardened exoskeleton of scleractinians, which are relatives of jellyfish within the phylum Cnidaria. These colonial organisms form rigid bodies anchored to the seafloor which form a complex three-dimensional structure offering shelter and feeding opportunities for many other species, so that reefs harbour one quarter of all modern marine species. These corals, filter feeders, also obtain part of their nutrition by symbiotically associating with algae of the zooxanthellae group (photosynthetic protists with a typically brown coloration). For this reason, they are forced to locate within the pelagic zone (<200m depth) where enough sunlight penetrates the ocean to support photosynthesis.
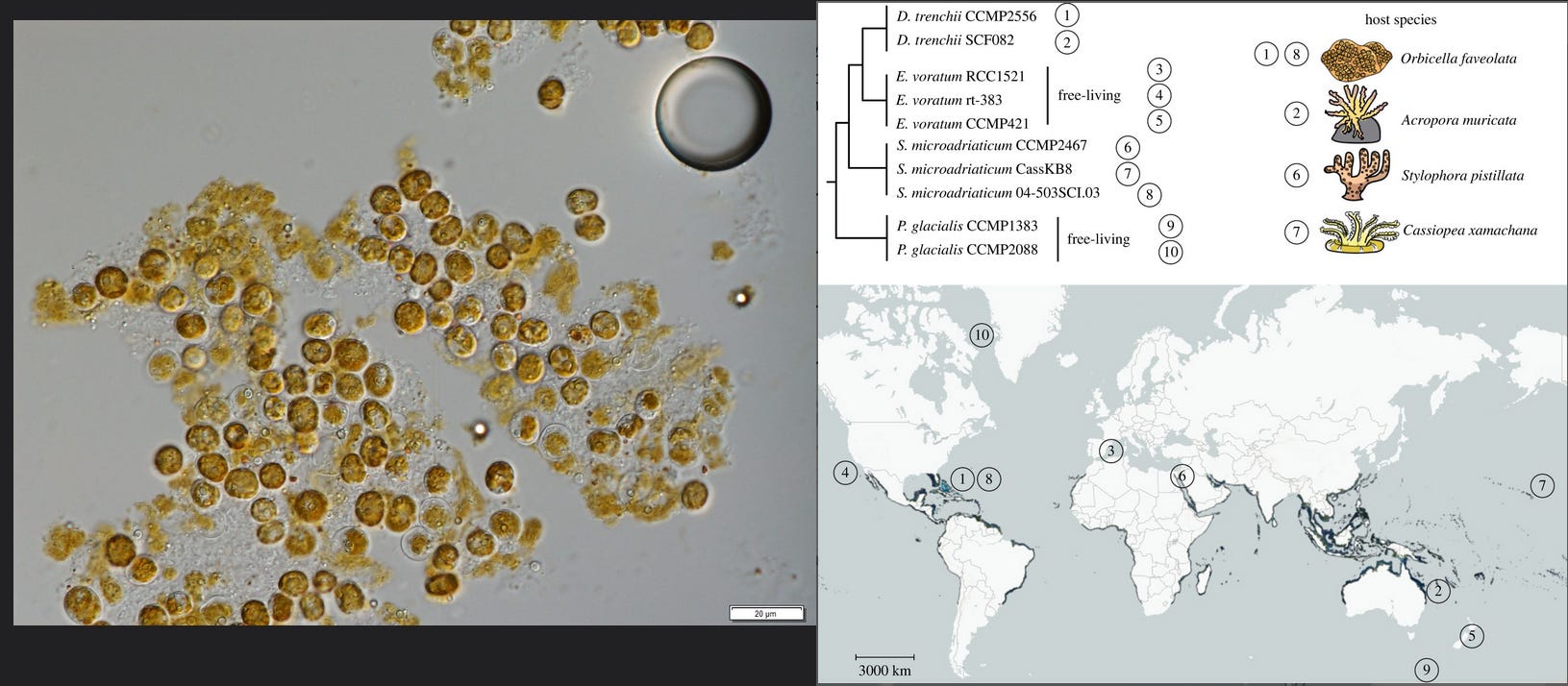
In a typical coral reef, there may be more zooxanthellae per m² than there are humans on Earth (10^10 zooxanthellae, for reference). Despite their number, ecological importance, and ubiquity in coral reefs, their biodiversity isn’t well understood, and the phylogenetic classification and population genetics are still contentious after decades of research, mainly because of the wide distribution of some species and the large genetic differences between populations widely considered to be conspecifics. Furthermore, they are not always strictly associated with a given host , but can be found associated with different species, sometimes even outside the scleractinians. It’s theorised that this lack of specificity is due to some corals not transmitting symbionts to their offspring, requiring them to form a new relationship with whatever compatible species are found living freely in the area.5
Surprisingly, despite what I said earlier about the fact that most ancient ecological niches have been filled in the sea long before they were on land, scleractinian coral reefs are the Rocky Balboa, underdog success story of the Cenozoic, having had variable fortunes over the course of their approximately 240 million year career as reef-building organisms. Starting as an insignificant (within the context of reefs) group of relatively uncertain origin in the mid-Triassic, they were nonetheless widespread and already differentiated into three distinct orders and six families, indicating that the earliest fossil record for this group may be fragmentary at best. After undergoing a remarkable radiation in the Jurassic, that saw them becoming dominant reef-builders, they were usurped in this role and they waned during the Tertiary. It wouldn’t be until the Eocene, some 50 million years ago, that they’d reclaim their spot as the premier reef builders of tropical shallow waters.67

This paragraph, of course, begs two questions. The first: if today’s scleractinians only became dominant reef-builders during the Jurassic, who built reefs before their first radiation? And secondly: how does a vital ecosystem engineer group “lose its job”, to end up replaced by someone else? This essay will delve into these two questions.
The dawn of the marine reef
Considering a reef as being a mineralised structure of organic origin found within the marine environment, the earliest contenders for the title may be some of the earliest fossils ever found, the Archaean stromatolites. Stromatolites are mineralised mounds composed of many distinct layers piled one on top of the other. The organic theory holds that microbial mats, while carrying out the chemical reactions needed to produce energy, caused the precipitation of compounds in shallow Archaean waters which led to the “seeding” of stromatolites with the formation of a rocky core on which subsequent mineral strata were deposited over the course of aeons.
Dating as far back as 3.4 billion years, these layered structures have long puzzled scientists. The two main issues were the following: are stromatolites organic in nature, or the result of geological processes, and, if organic, how were they produced over a billion years before the rise of cyanobacteria and the great oxygenation event 2.5 billion years ago? A 2016 paper proposes theories as to their formation.8

To simplify a relatively in-depth biochemical explanation, it’s believed that microbial photosynthesis existed long before the oxygenation event, divided between two distinctive anoxic modes that varied depending on environmental conditions thanks to a molecular “switch” that activated one or the other. The first pathway would have used, instead of water, inorganic molecules like H2S or ferrous iron as electron donors, while the second used organic molecules such as pyruvate.
It is likely that hydrothermal activity, still prevalent on this young, turbulent Earth, would have introduced large amounts of inorganic electron donors to certain areas, and it’s in the shallow, photic zones near these hydrothermal vents that microbial mats would have formed by harnessing the free energy of the Sun using an alternating supply of inorganic and organic molecules as electron donors, as conditions dictated.
As rainfall, tidal activity or the vagaries of hydrothermal activities increased or diluted H2S concentration, the microbial mats switched to one of the two photosynthetic pathways, each switch leading to an accumulation of distinctive compounds and trapped sediment over time layering on top of one another. This also had the advantage of bringing each new layer closer to the surface, increasing photosynthetic activity.
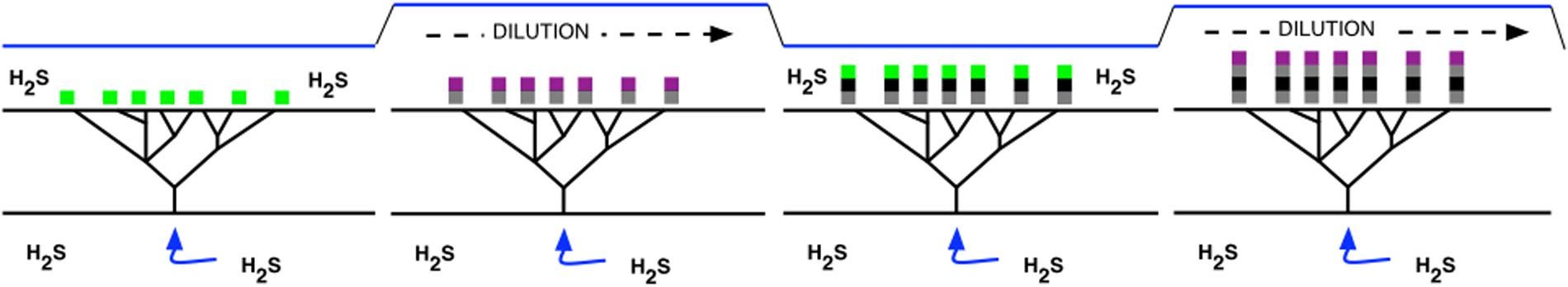
Despite this unfathomable antiquity, stromatolites still exist today, although the microbes which cause them are varied and not necessarily considered to be related to their Archaean-era predecessors (as typically, they’re initiated by oxygen-forming cyanobacteria now). They’re most well known from extreme environments such as hypersaline lagoons or thermal springs, but can also thrive in other biomes.

The first animal-built reefs
After many aeons of not much happening on this front, the reef-building niche would take its first step forwards. New, multicellular, organisms would lead to the creation of new reef structures in the oceans. This took place during the Ediacaran, some 548 million years ago, even before the Cambrian explosion which gave birth to complex marine ecosystems. The early Metazoans responsible for this were a genus called Cloudina, the earliest known Nephrozoans (ancestors of all living animals except for Cnidarians like jellyfish and coral, ironically, and a few other odds and ends). These minuscule tube-like organisms secreted a series of imbricated calcareous cones. Their ecological preferences and lifestyle are largely unknown.
These organisms attached themselves to stromatolites near shorelines and orientated themselves in a consistent way throughout the community, in reefs sometimes kilometres long. It’s currently supposed that they were filter-feeders, attaching to an elevated solid substrate and to each other in order to best reach the food-rich currents they depended on for survival. Like all good reefs, this also created attachment points for various other gregarious sedentary organisms such as Namapoikia or Namacalathus.9

The most famous early reef-building animal is the lower-Cambrian Archaeocyatha. An early type of sea-sponge. Like those of today, ancient sea-sponges were sessile animals that fixed to a hard substrate and lived by filtering the water. They generally formed relatively small reefs although this varied locally. While their reefs could be found all throughout the Cambrian, they disappeared at the end of that geological era, leading to a long gap with few animal reefs being built.10
The first coral age
While reefs up to this point were still dominated by microbes, they would take a new direction during the Ordovician, following the Great Ordovician Biodiversity Event (GOBE). Stromatolites would give way to true coral reefs composed of a combination of a new group of sea-sponges called Stromatoporids, and the very first coral reef-builders, two different groups known as rugose and tabulate corals.
The primarily Bryozoan (sea-sponge) based reefs of the early Ordovician were all dome-shaped, but varied in size from structures comparable to a dinner plate, to some several metres wide and as tall as a person.11 It must be noted that, based on other factors similar to those discussed below, they’re considered to have potentially been photosymbiotic, like modern coral and sea-sponges. Stromatoporids, ultra-calcified sea-sponges, would become the main reef-builders of the Ordovician and Devonian, upon which the smaller colonial corals would develop.12

The tabulates appeared first in the lower Ordovician. Some of their fossils are easily recognisable as they formed massifs of linked tubes that, from above, look like the links of a chain. They formed heavily integrated colonies whose individuals rarely measured more than a few millimetres in width but, unlike modern Scleractinian corals, secreted the calcium carbonate of their tough exoskeletons in the form of calcite, as opposed to aragonite. They rarely formed the basis of reefs, preferring to attach on pre-existing solid structures. Recently, it was determined that most tabulates were probably photosymbiotic like modern Scleractinians, based on the presence of seasonal growth bands, the dense and highly integrated colonial structure, and measures of isotopic ratios of oxygen and carbon within the calcite structure. Since Symbiodiniaceae algeae only appeared during the Eocene, another group of single-celled photosynthetic organism must have been the basis of this association. However some deep-sea species from below the photic zone had asymbiotic isotopic markers.13
The rugosans meanwhile are from the middle Ordovician. Their main characteristic was their horn-shaped calcite skeleton, with a rough outer texture that would give them their name. It’s believed they have evolved independently from an ancestral non-skeletal anemone.14 These corals, sometimes solitary and sometimes reef-building, were always subordinate in terms of biomass to the tabulates, and although the two could coexist, the fossil record indicates that they had different ecological preferences and likely did not compete with each other.
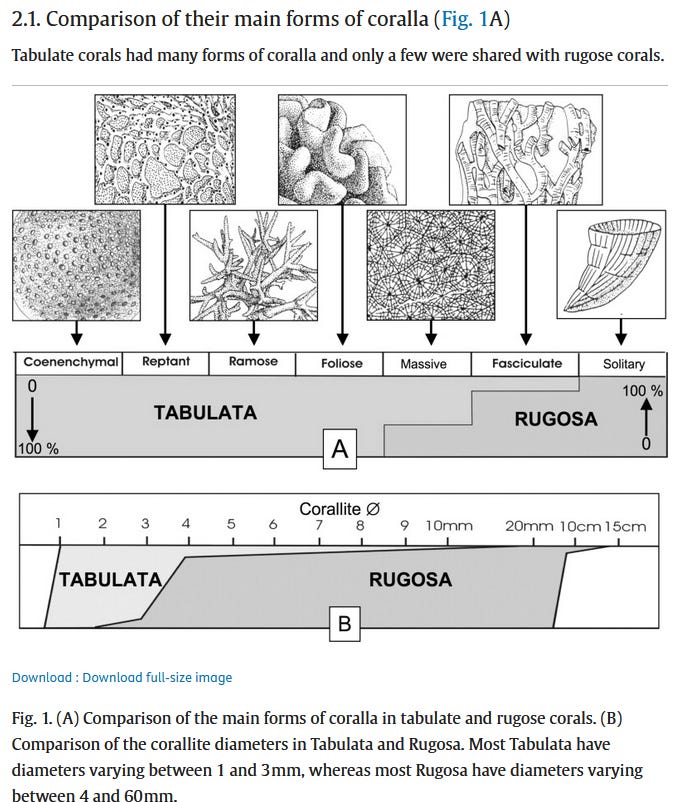
Both coral groups would last through several geological epochs, surviving a variety of mass extinctions and other ecological crises. The tabulates, however, would never recuperate from the Frasnian event, a mass extinction which took place at the end of the Devonian, and disappear from the fossil record along with the stromatoporids at this time. The rugose corals meanwhile would “limp on” with diminished morphological diversity until the Permian-Trias extinction. 15
The early Mesozoic - it’s over, we’re back
If there’s one term that can describe the fortunes of reef-building organisms after the Permian-Trias extinction, it’s certainly “boom and bust”. The extinction was the most severe one of the Phanerozoic, eliminating between 80-96% of all marine species. This event hit sessile filter feeders especially hard, leading to the proliferation of mobile organisms like gasteropods in the aftermath. After a long interval of minimal reef activity, caused by a conjunction of factors including ocean acidification, a few “holdover” organisms from the Permian would start building sizeable reefs again in the Mid-Triassic. This activity would be centered around the Tethys ocean and involve Permian bryozoans retaking their place as main reef builders. However, this return would be brief, as the late Triassic extinction would eliminate all but one clade of bryozoans. 16 As a footnote, during the Triassic, a strange group of tiny organisms of uncertain taxonomic origin appears to play a major role in reef formation. Known as Tubiphytes, they have been compared to everything from cyanobacteria to protists, but some research argues they are related to sea-sponges.17
Following a major reshuffling of marine communities, the Jurassic is described as a period of transition for marine reefs. In the mid to late Triassic, an unknown group appears in the reefs of the Tethys ocean, clearly differentiated from pre-existing coral groups, and rises to prominence following their extinction. These would be the first Scleractinians, ancestors of all modern corals, believed to have been derived from soft-bodied sea anemonies which independently developed calcium carbonate skeletons (an explanation for the reason why they appear suddenly in the fossil record fully differentiated in several families). 18 As I’ve already explained earlier, they would enjoy a period of prominence starting from the late Triassic until the Cretaceous before being “usurped” in their ecological role.
That’s quite rude(ist)
After the late Triassic extinctions, which once again greatly reduced reef diversity and distribution, Scleractinians would restart colonising the suitable seashores of the world over the course of the Jurassic. After a brief boom in the late Jurassic, an unexpected, dark-horse contender suddenly monopolised this ecological niche.
These upstarts would appear in the fossil record during the mid-Jurassic inside the Tethys sea, which would much later become part of Europe. Dubbed “rudists” by Lamarck because of their rough texture, the numerous horn-shaped fossils spent many years being switched between different animal phyla, until scientists finally settled on classifying them as bivalves. They thrived in the warmest tropical waters near the equator, spreading much like modern bivalves thanks to their zooplanktonic larvae being carried by the dominant sea-currents. As the currents favoured transportation from the “Atlantic” half of Tethys to the “Caribbean” half, with fewer possibilities of return, the populations of these two regions speciated. Rudists are, to this day, the only bivalves to have been the world’s dominant reef-builders. They were filter-feeders, like most bivalves known today, and indeed like most reef-building organisms.

By the time of the middle Cretaceous, rudists in the “Caribbean” region had grown in numbers until they crowded scleractinians out of the top spot in terms of biomass on reef ecosystems. The “Atlantic” ones would follow a few million years later. They were among the largest bivalves ever recorded, some measuring over a metre in length. Unlike most modern bivalves, they could be extremely asymetrical, the largest species needing to “stand” upright in the sediment and congregating to support each other against the current, with their second valve being a flat flapper which opened and closed to protect the animal or allowed it to feed. These reefs could run for hundred of miles continuously, parallel to the shores, and be hundreds of metres tall, build on layers of accumulated shell debris cemented together with sediment.19
While rudists would survive several mass extinction events over the course of the Jurassic and Cretaceous, they would ultimately disappear during the infamous K/T extinction event which also saw the extinction of the dinosaurs. It must be noted however that a decline in their diversity was already taking place a few million years before then.20

The Cenozoic: return of the king, and other contenders
The downfall of the rudists brings us to the final chapter of our journey through time, as scleractinians would eventually emerge from their refugia deeper in the sea to reclaim the shorelines of the world during the Eocene, ushering in the modern era of reef-building which contains the largest surface of scleractinian reefs of all time.
Throughout these historical vignettes, we’ve seen major reef-builders rise from insignificant species and take over, only to vanish in turn, to be replaced by another group. In all cases, after the Archaean, reefs were never mono-specific affairs, or even built by a single clade. Many organisms throughout the tree of life contributed to the solid structure of the reef in all epochs through convergent evolution, and some of them are still alive today.
While scleractinians have proven uniquely resilient against mass extinctions, repeatedly recuperating from events which annihilated most of their contemporary fauna, it may be interesting to look at a few of the other reef-building species alive today and imagine what would happen if corals were indeed wiped out, or if these upstarts developed strategies allowing them to become the dominant reef-builders of the future.
Vermetids are marine gasteropods (“snails”) with a peculiar morphology involving the secretion of a tube-shaped calcium carbonate shelter that they never leave, only interacting with the outside world by opening an airtight flap at the summit in order to filter feed. They have extremely low dispersive power, as male sperm is released as spermatophores into the water where it has to quickly be collected by a feeding female. She’ll harbour it until water temperatures allow fertilisation and brood the young inside her mantle. These crawling juveniles only travel for a few hours at most before starting to secrete their own tube, which they’ll never leave. This leads to the group having very high diversity between populations and many cryptic species. They preferentially form reefs in warm waters, with their largest populations being centered in the Caribbean and Mediterranean seas. They’re threatened by ocean acidification and habitat destruction.21

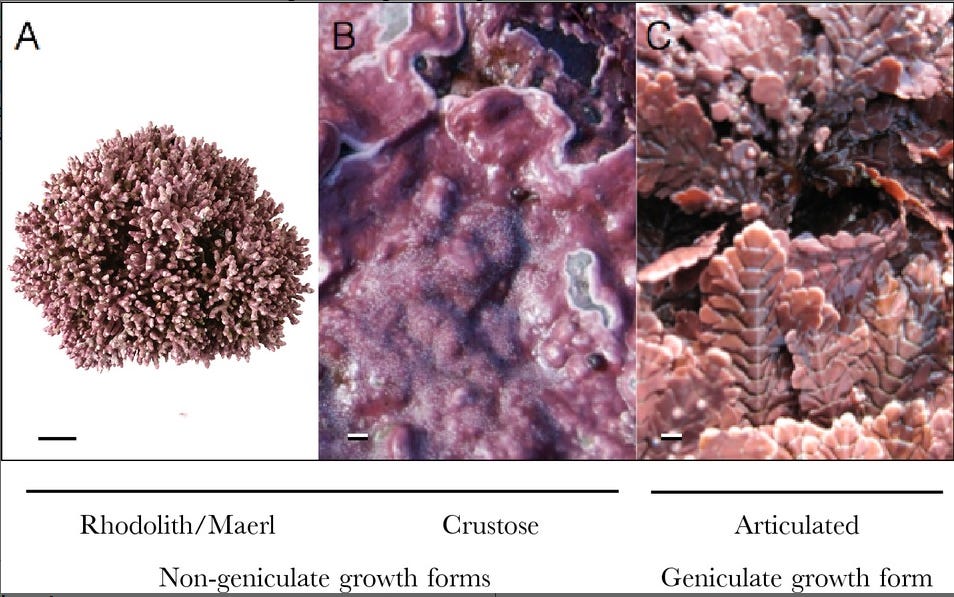
Closeup of a Dendropoma cristatum vermetid reef, by Frédéric André. Coralline algae are part of the “red algae” clade, so-called because of their peculiar pinkish-red coloration caused by the presence of specific photopigments. They have the peculiarity of secreting calcium carbonate in their cell walls, giving them a tough, rigid structure. They are mainly divided into non-geniculate and geniculate groups. The non-geniculate coralline algae do not have articulations in their exoskeleton, meaning they grow in fine straight filaments which focus on vertical growth to reach the sunlight. They may grow as fast as 1cm per year. Geniculate forms meanwhile have articulations which give them greater flexibility, frond surface and photosynthetic capacity, allowing them to grow three times as quickly. These algae may be the primary reef builders in some regions, and generally provide an important contribution to coral reefs worldwide by “cementing” the coral debris together into a coherent structure. 22

{kind=link}
{kind=link}
Serpulid worms are small (<2cm) marine annelids which secrete a calcium carbonate tube attached to a solid substrate, from which they’ll feed by filtering the surrounding water using an impressive array of fan-like structures. In rare circumstances, millions of individuals will congregate in a given environment, forming reefs similar to those of vermetid snails that provide a complex 3D structure in which small fauna may shelter and brows for food, enriching the area’s biodiversity. Baffin Bay, in Texas, is an example of this. These worms typically endure environments too saline for other filter feeders.23

{kind=link}
While rudists are the only bivalves who became the world’s top reef-builders for a time, they’re not the only ones to still build reefs today. Shellfish reefs are generally found in colder waters unable to support coral growth, and provide many ecological services, not least of which are water purification through filter-feeding and management of algal blooms, while also increasing biodiversity.24 They are generally composed of species commonly called “mussels” or “oysters”. Australia’s largest marine restauration project involves the re-creation of the extensive oyster reefs of its Southern coastline, eradicated by the fishing and lime industries during the 19th century.25

Conclusion
An ecological niche, much like a species, is something that only makes sense within a specific time and place. For this reason, niches come and go over geological time, their occupants becoming dominant or exiled depending on how easy it is to thrive in that given lifestyle. Even reef-building, one of the oldest niches in existence, went through regular upheavals over the course of its 3.5 billion year history, and doubtless will continue to do so. Although scleractinians have displayed exemplary staying power over the last 240 million years, how long are they going to keep going, and who will replace them? It’s certain that we won’t know within our lifetimes, and maybe even the lifetimes of all of Mankind. But it’ll certainly happen some day, and I hope this post gave you a few ideas how that might go.
1 Sousa et al., 2008. Ecology of the invasive Asian clam Corbicula fluminea (Müller, 1774) in aquatic ecosystems: an overview. https://www3.epa.gov/region1/npdes/merrimackstation/pdfs/ar/AR-1406.pdf
2 https://www.nature.com/scitable/blog/accumulating-glitches/the_first_forests/
3 https://www.scientificamerican.com/article/revealed-the-first-flower-140-million-years-old-looked-like-a-magnolia/
4 Khramov et al., 2023. The earliest pollen-loaded insects from the Lower Permian of Russia The earliest pollen-loaded insects from the Lower Permian of Russia
5 Baker, 2018. Flexibility and specificity in coral-algal symbiosis: diversity, ecology, and biogeography of Symbiodinium. doi:10.1146/annurev.ecolsys.34.011802.132417
6 Stanley, 1981. Early history of scleractinian corals and its geological consequences. GEOLOGY, v. 9, p. 507-511
7 Stolarski et al., 2011. The ancient evolutionary origins of Scleractinia revealed by azooxanthellate corals. https://doi.org/10.1186/1471-2148-11-316
8 Allen, 2016. A Proposal for Formation of Archaean Stromatolites before the Advent of Oxygenic Photosynthesis. https://doi.org/10.3389/fmicb.2016.01784
9 Penny et al., 2014. Ediacaran metazoan reefs from the Nama Group, Namibia. https://doi.org/10.1126/science.1253393
10 Pruss et al., 2021. Cambrian reefs in the lower Poleta Formation: a new occurrence of a thick archaeocyathan reef near Gold Point, Nevada, USA. https://doi.org/10.1007/s10347-021-00623-2
11 Adachi et al., 2011. EARLY ORDOVICIAN SHIFT IN REEF CONSTRUCTION FROM MICROBIAL TO METAZOAN REEFS . https://doi.org/10.2110/palo.2010.p10-097r
12 Hong et al., 2017. Construction of the earliest stromatoporoid framework: Labechiid reefs from the Middle Ordovician of Korea. https://doi.org/10.1016/j.palaeo.2017.01.017
13 Zapalski, 2014. Evidence of photosymbiosis in Palaeozoic tabulate corals. https://doi.org/10.1098/rspb.2013.2663
14 Baars et al., 2013. The earliest rugose coral. doi:10.1017/S0016756812000829
15 Poty, 2010. Morphological limits to diversification of the rugose and tabulate corals. https://doi.org/10.1016/j.palwor.2010.09.006
16 Powers et al., 2008. DIVERSITY AND DISTRIBUTION OF TRIASSIC BRYOZOANS IN THE AFTERMATH OF THE END-PERMIAN MASS EXTINCTION. https://www.jstor.org/stable/20144200
17 RIDING, R., GUO, L. 1992. Affinity of Tubiphytes. Palaeontology, 35, 1, 37–49. https://www.palass.org/publications/palaeontology-journal/archive/35/1/article_pp37-49
18 Stanley. 1988. The History of Early Mesozoic Reef Communities: A Three-Step Process. https://doi.org/10.2307/3514528
19 Johnson, 2002. The Rise and Fall of Rudist Reefs: Reefs of the dinosaur era were dominated not by corals but by odd mollusks, which died off at the end of the Cretaceous from causes yet to be discovered. https://www.jstor.org/stable/27857629
20 https://ucmp.berkeley.edu/taxa/inverts/mollusca/rudists.php
21 Milazzo et al, 2016. Drawing the Line at Neglected Marine Ecosystems: Ecology of Vermetid Reefs in a Changing Ocean. DOI 10.1007/978-3-319-17001-5_9-1
22 Mccoy et al., 2014. Coralline algae (Rhodophyta) in a changing world: integrating ecological, physiological, and geochemical responses to global change. https://doi.org/10.1111/jpy.12262
23 Palmer et al., 2021. Importance of Serpulid Reef to the Functioning of a Hypersaline Estuary. https://doi.org/10.1007/s12237-021-00989-0
24 Christianen et al., 2017. Biodiversity and food web indicators of community recovery in intertidal shellfish reefs. https://doi.org/10.1016/j.biocon.2016.09.028
25 https://www.natureaustralia.org.au/what-we-do/our-priorities/oceans/ocean-stories/restoring-shellfish-reefs/