


Oldies but goodies
Crash course in angiosperm taxonomy and reproductive anatomy

I recently responded to a viral post of the “nature is amazing” kind about a peculiar-looking genus of flowering plants, Dendrosenecio, found in African alpine environments (such as the Kilimanjaro). I’ve always found them interesting as an example of radiative adaptation, which can be guessed at by looking at the genus name. “Dendro” is derived from a Greek word meaning “tree”, while Senecio is the name of a very widespread and diversified genus of small-to-middling-sized herbaceous plants, often considered to be weeds (although this genus was recently split off of Jacobaea, taking a goodly chunk of European species with it, which is a whole other story). This isn’t a coincidence, since the genus is a divergent splinter of Senecio shaped by the extremely hostile conditions faced by plants living in altitude.

What had struck me about the post was the use of the word “prehistoric” to describe the plants. Indeed, they have a very peculiar body plan compared to most temperate plants, with stout, thickened trunks sheathed in dried out leaves more reminiscent of palm trees, or tree-ferns, than of modern flowering plants. Although many were quick to point out the more general issue that the word “prehistoric” makes no sense in the context of describing wildlife, since recorded history only began a few millennia ago, I believe the word here was more used to mean “pre-cretaceous extinction”. Indeed, it’d be easy to imagine an allosaurus lurking within a Dendrosenecio forest.

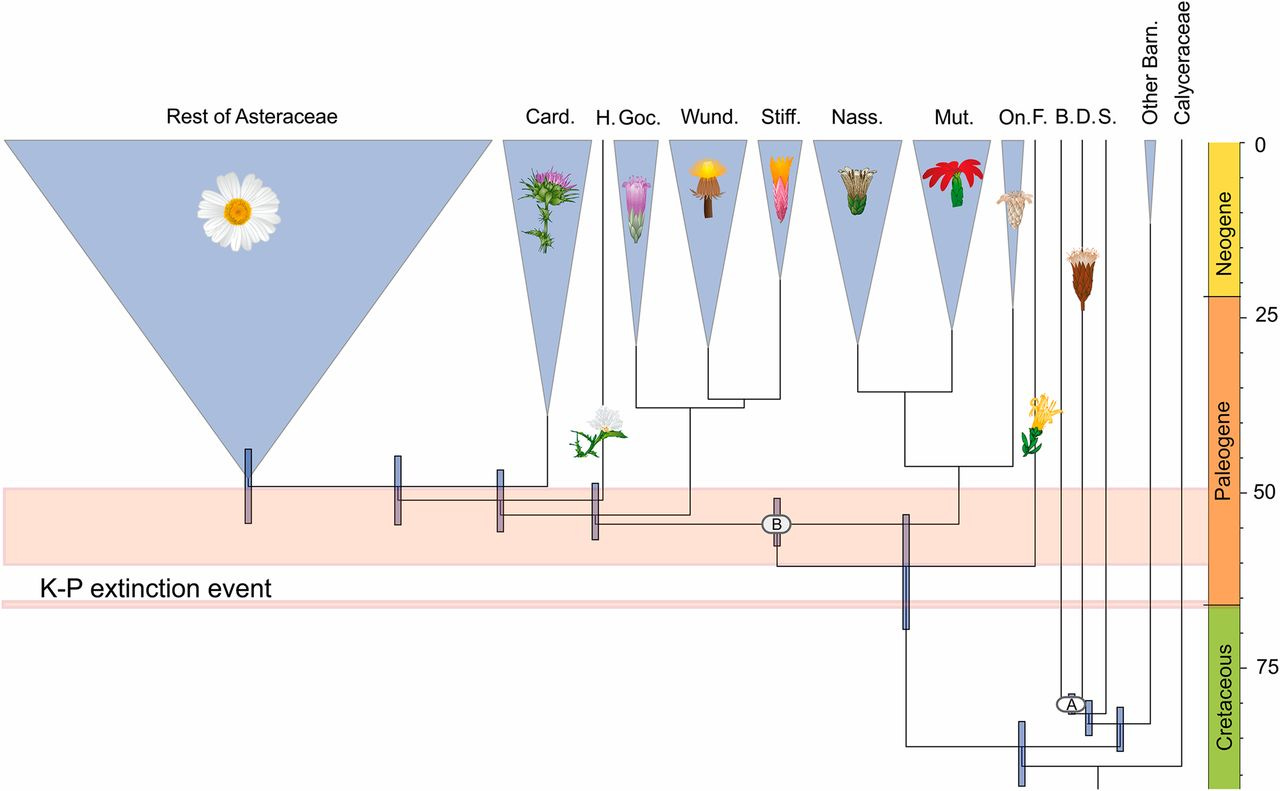
The interesting tension here was that evolutionarily speaking, the Asteraceae is not a very old family chronologically, and is also extremely derived morphologically from the layout of the earliest Angiosperms (flowering plants). To put it into concrete numbers, the tribe Dendrosenecio belongs to, Senecioneae, differentiated with most of the rest of its family during the Paleogene, around 45-50 million years ago (the family itself appears in the late Cretaceous).1 Within this tribe, the genus Dendrosenecio only split from Senecio within the last million years, actually making it quite young by plant standards. For contrast, the rose family Rosaceae, which few laypeople would consider particularly primitive-looking, was recently found by phylogenetic analyses to be approximately 120 million years old2, with the genus Rosa already appearing in the fossil record 55 million years ago.
What am I getting at?
While talking about plant phylogeny by itself is extremely interesting to me, I had a more practical point to make about botany. Since plants are a whole other kingdom of live, we aren’t attuned to their morphological peculiarities and variations in the same way as we are for vertebrates, so it’s not always easy for a regular person to tell which plants fit together and what are common ancestral traits of plants, which were lost or kept over geological time scales. It should be noted that with plants, there is far more plasticity in the vegetative parts than the reproductive organs (flowers and fruits), since any even slightly maladaptive floral traits will have a far more immediate effect on an organism’s viability in evolutive timescales than anything to do with how the plant grows (beyond of course vegetative traits causing quick death). This means that floral traits and fruit morphology are more likely to be conserved within clades of plants, while foliage can vary wildly
We can tell reconstructions of premodern humans have more simian traits such as the small cranial volume, the shape of their forehead, the size of their jaw and how bipedal they are, but how to tell if a plant has very conserved basal traits or not? I’ll list the most basal traits found in ancestral angiosperms and list a few examples of plants that have them.
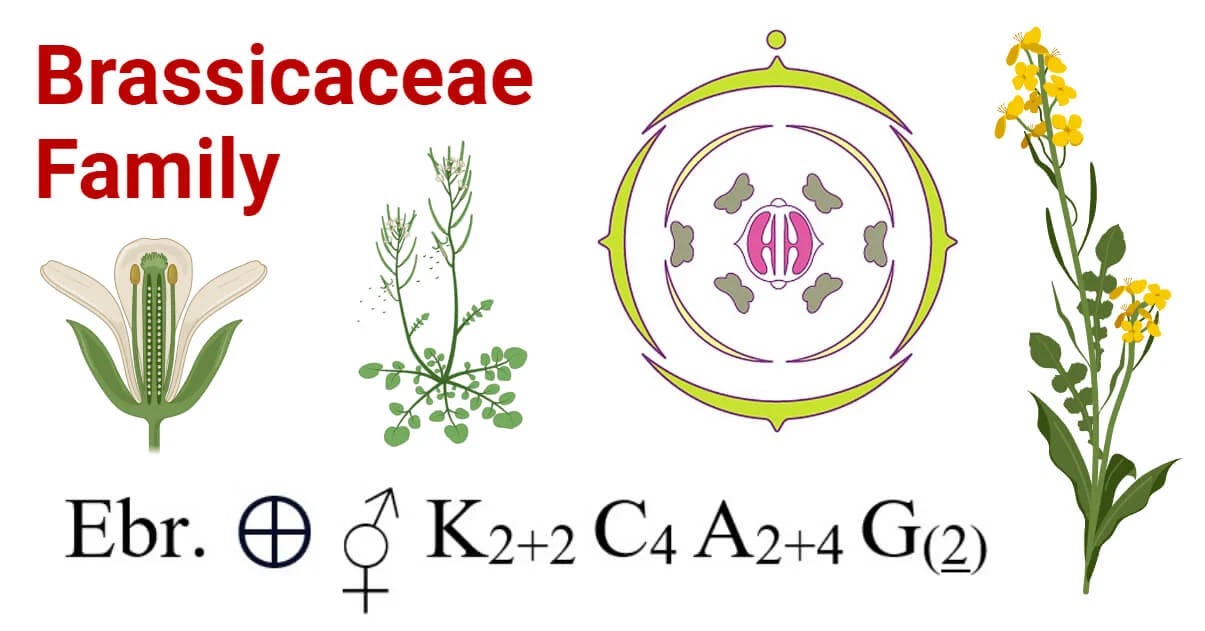
As an aside, there is a standardised and condensed nomenclature used for describing the structure of a flower scientifically, called the floral formula. I never really had to use it, although I have a couple books that do. I think it’s mostly useful if you’re a professional plant taxonomist or someone who goes out there to look for and describe new species. Here’s what it looks like in practice (I won’t be using it but that way you know it exists).
The first flowers
A 2017 paper published in Nature purported to model the flower of the earliest known ancestor of all currently living Angiosperms, by sampling the floral traits of 792 species from 98% of all angiosperm families and calibrating their molecular dating analysis with 136 fossils.3 I’m fairly sure I’ve mentioned it in a post somewhere before, but can’t remember where or why.
Their resulting “most basal flower” has several distinctive traits one can still find in some plants today: it is bisexual, radially symetrical, has numerous (more than 10) free stamens (male organs) and more than five carpels (female organs). Meanwhile, its (numerous, more than ten) perianth organs are undifferenciated (it has a single set of colourful floral elements called “tepals”, as opposed to a set of petals and sub-tending sepals). All the organs were free from each other (no fusion takes place during flower formation) and the carpels were superior (resting on top of the floral receptable rather than integrated into it and protected by it). What does that look like in practice?
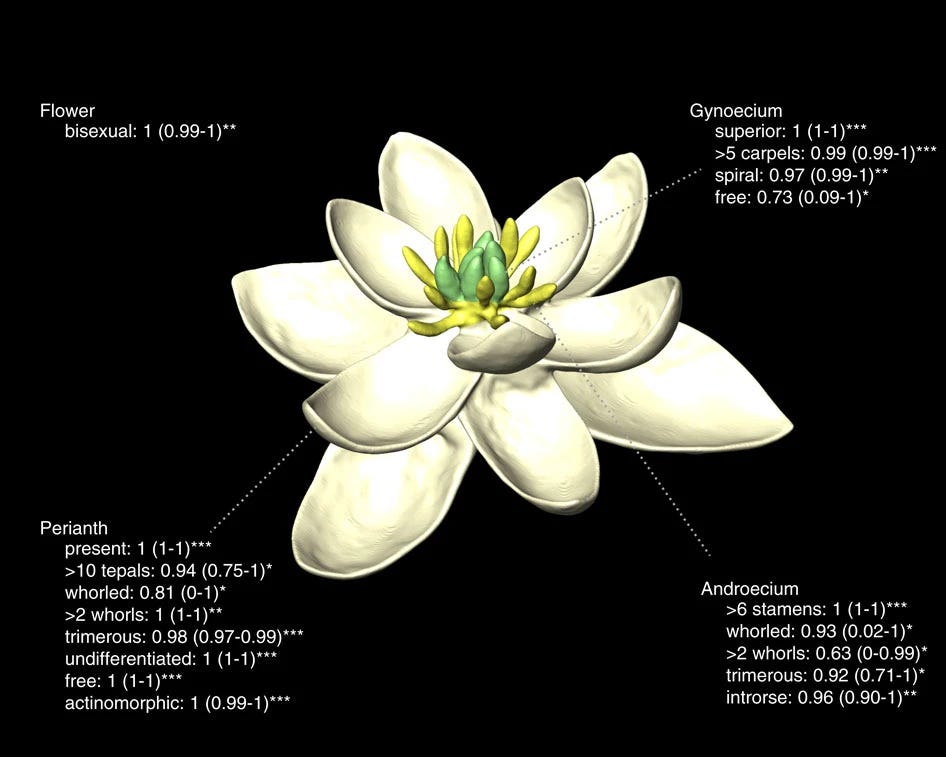
Those of you who visit parks or have a particularly large garden might find this flower somewhat familiar, as it looks essentially identical to those found in the genus Magnolia, very frequently planted as an ornamental in temperate climates. This could be because, as botanists have long known, magnolias are one of the groups that have the most conserved basal traits among Angiosperms, or else it could be that this belief lead to a bias in the interpretation of the data, leading to a very similar product. Most noticeable are the prevalence of unfused elements, numerous undifferentiated tepals, radial symetry, and a (very!) large number of free stamens (still immature in this picture, at the base of the middle of the flower) and carpels. For reference, recognisable magnolia plants enter the record 95 million years ago.4

Other species with a similar floral layout include water-lilies (such as Nuphar lutea) which are also known to phylogenists as one of the basal flowering plant families but also the hellebores (Helleborus viridis for example, which I wrote about here in the past). These latter plants are some of the most “basal” looking among their family Ranunculaceae (which is also an ancient family), but it should be noted that some members of the family do have differentiated petals and sepals, or even bilateral symetry, so there is more variation.
On the opposite end of the spectrum are plants with extremely transformed flowers, often presenting extensive fusion between various organs, and even fusion between numerous flowers into a single structure, the stunning tropical plant Strelitzia is a fine example of this. Its inflorescence is sheathed within a colourful beak-shaped bract (modified leaf sub-tending a flower) and contains 4-6 flowers, each comprised of a pair of showy structures. The blue ones are petals fused with stamens (themselves fused into a tube-shape to fit into the petal-cone) to ensure pollination, the stigma is also fused nearby for ease of access, while the orange ones growing around the blue ones contain pools of nectar.

Then you have everything else between these two extremes. Brassicaceae which have a few free petals and also few anthers, Caryophyllaceae which have very differentiated sepals and petals and often relatively few carpels, Araceae with their extremely transformed, radar-dish-like inflorescences, etc etc.
Where does this leave Dendrosenecio?
Being a close relative of the genus Senecio, it is of course a member of the family Asteraceae, the composite family. They are so-called because each of their “flowers” is actually a highly transformed and specialised conglomerate of minuscule flowers, which together imitate a single larger flower. To take the example of the common daisy (Bellis perennis):

Asteraceae often have two types of flowers, peripheral “ray florets”, which have a bilateral symetry, as part or all of their petals fused into a single long sheath called a “ligule” (this imitates the petals of non-composite flower types) while the innermost ones (“disc florets”) usually have four or five fused petals and a radial symetry. This extensively modified arrangement is a far-cry from the basal flower types we’ve seen above! Now let’s take a look at the flower of Dendrosenecio specifically.

{kind=link}
We see that, despite its outlandish vegetative body, and extreme adaptations to its hostile environment, Dendrosenecio is at heart a “classical” Asteraceae, its flowers morphologically identical in all respects to those of its brethren within the Senecioneae tribe. Thus, from a morphological standpoint, it is far from being part of the groups of flowers which we would rightly call “basal”, or to put it in cruder terms “primitive” or even “prehistoric”.
Conclusion
When studying organisms so different from us, we must avoid the shortcuts and generalisations we use to study our closest relatives the vertebrates. When it comes to plants, their sexual organs and fruits are often (but not always!) the best way to identify kinship and discriminate between similar species. But this also means that kinship can be established between even the most outlandishly different plants using the reproductive organs, as this morphologically is often the most conserved part of their anatomy. Thus, studying plants requires adapting your mindset to the evolutionary logic of a lineage that’s been separate from ours for most of life’s existence on Earth.

Barreda et al., 2015. https://www.pnas.org/doi/full/10.1073/pnas.1423653112
Caspermeyer, 2017. https://academic.oup.com/mbe/article/34/2/505/2966070
Sauquet et al., 2017. https://www.nature.com/articles/ncomms16047
Strelitzia is commonly known as “bird of paradise”. This is a wonderful article which I read because Stone Age Herbalist alerted me to its existence!